
Neutrální model evoluce a nedávné počátky v Africe
Autor: dr. Robert W. Carter
V originále vydáno: 2. července 2010
„Výzvou genetických studií historie lidstva je využít malého množství genetických diferencí mezi populacemi k odvození historie lidských migrací“ (Rosenberg et al. 20021).
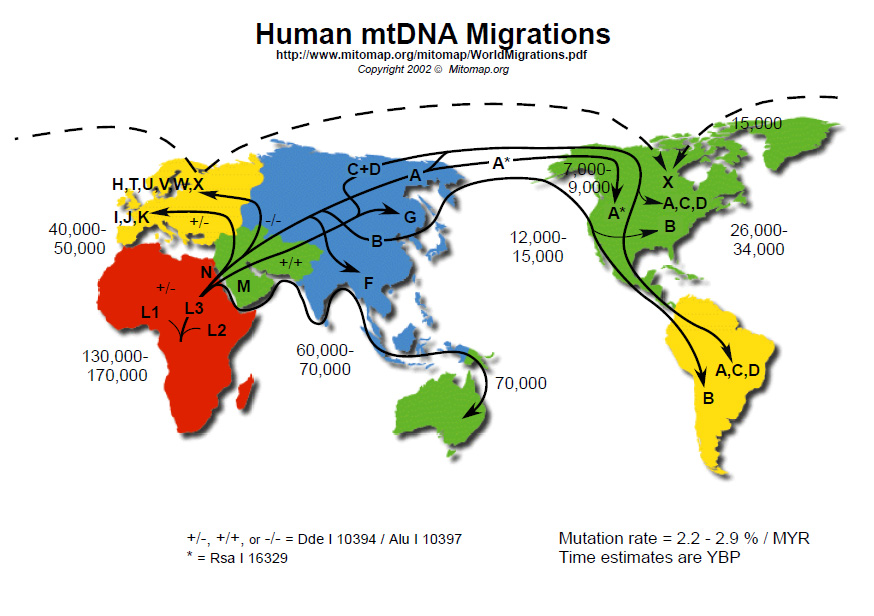 Obrázek 1. Tato mapa podrobně zobrazuje koncept nedávného afrického původu. Podle teorie RAO vznikli moderní lidé v Africe, tam se diverzifikovali a pak jedna linie (s několika hlavními klady) migrovala pryč, aby osídlila zbytek světa. Je to velmi podobné biblickému pojetí, jen s jiným výchozím bodem (Blízký východ) a jiným časovým rámcem. (Převzato z MITOMAP: A Human Mitochondrial Genome Database (Databáze lidského mitochondriálního genomu), <www.mitomap.org>, 2008). Klikněte sem pro větší zobrazení. Obrázek 1. Tato mapa podrobně zobrazuje koncept nedávného afrického původu. Podle teorie RAO vznikli moderní lidé v Africe, tam se diverzifikovali a pak jedna linie (s několika hlavními klady) migrovala pryč, aby osídlila zbytek světa. Je to velmi podobné biblickému pojetí, jen s jiným výchozím bodem (Blízký východ) a jiným časovým rámcem. (Převzato z MITOMAP: A Human Mitochondrial Genome Database (Databáze lidského mitochondriálního genomu), <www.mitomap.org>, 2008). Klikněte sem pro větší zobrazení.
|
Tento citát ilustruje jednu důležitou věc. Moderní genetici se snaží pochopit genetickou historii člověka. Nakonec jsou nuceni učinit určité závěry na základě omezených údajů a řady zjednodušujících předpokladů. Účelem tohoto článku je podívat se na základy modelu nedávného afrického původu (RAO) lidské evoluce, který byl poprvé zpopularizován tvrzením o objevu „mitochondriální Evy“ v Africe.2 Každý ze základních předpokladů této teorie byl v evoluční literatuře otevřeně zpochybněn. Pokud některý z předpokladů RAO padne, může se celá teorie stát bezpředmětnou. Doufám, že výčtem předpokladů a následným systematickým ukazováním, jak je každý z nich nepraktický, nemožný, rozporuplný nebo neobjektivní z hlediska evoluční teorie, se mi podaří význam RAO o několik stupňů snížit.
Termín „nedávný“ používají zastánci RAO ve smyslu hlubokého času a nemá se chápat jako podpora mladé Země. Podle RAO se lidé vyvinuli v Africe, existovali jako malá populace po dobu několika set tisíc let a poté se zhruba před 200 000 lety rychle rozšířili do zbytku světa. Jako vysvětlující nástroj je přímým protikladem biblického modelu, kde by nejdůležitějšími genetickými signály měly být Stvoření (které omezuje celkovou lidskou rozmanitost), Potopa (událost hrdla lahve) a událost babylonské věže (která vedla k významnému dělení populace a celosvětové migraci). Očekává se, že poslední dva modely přinesou podobné výsledky jako hypotetický model RAO, ale v jiném časovém rámci.
Na tomto místě chci čtenáře varovat. Mohlo by se zdát, že tento článek vyvrací všechny argumenty založené na neutrální evoluci nebo teorii hrdla lahve (oba pojmy budou definovány níže). Kreacionisté tyto teorie někdy využívají ve svůj prospěch a mým záměrem není je zcela znemožnit. RAO ve skutečnosti používá stejnou matematiku, kterou by mnozí kreacionisté rádi použili na biblický model. Mým záměrem je zaujmout spíše „chirurgický“ přístup, vyříznout „rakovinu“ špatné vědy a ponechat nedotčenou vědu, která může být ještě platná a užitečná. Svou argumentací proti RAO vlastně obhajuji poměrně nedávný původ lidstva, ačkoli se zde na toto téma nezaměřuji a v tomto článku se nijak výrazně neprojeví.
„Cílem tohoto článku je poukázat na místa, která stojí na nerealistických předpokladech v jejich prospěch, a doufám, že poukázáním na nereálnost těchto předpokladů se mi podaří rozptýlit některé mýty modelu RAO [Recent African Origins].“
RAO se dopouští řady aproximací, stejně jako všechny teorie. Z tohoto důvodu může být téměř každá teorie napadena jako „nerealistická“. Cílem tohoto článku je poukázat na místa, která stojí na nerealistických předpokladech v jejich prospěch, a doufám, že poukázáním na nereálnost těchto předpokladů se mi podaří rozptýlit některé mýty modelu RAO.
RAO nepodporuje rozsáhlou evoluci v hlubokém čase. Na tom, že se lidé někdy v minulosti přestěhovali z Afriky nebo do ní, není nic evolučního. Význam této teorie spočívá spíše v otázce datování a ukotvení lidského rodokmenu. Následující citát ze zásadního dokumentu RAO bude středobodem všeho, co bude následovat: „Z toho usuzujeme, že Afrika je pravděpodobným zdrojem lidského mitochondriálního genofondu. Tento závěr vyplývá z pozorování, že jedna ze dvou primárních větví vede výhradně k africké mtDNA (mitochondriální DNA)…, zatímco druhá primární větev vede také k africké mtDNA.“3 Podaří-li se nám toto analyzovat, výrazně pokročíme na cestě k lepší teorii genetické historie člověka (obrázek 1).
Standardní neutrální model evoluce
RAO vychází ze standardního neutrálního modelu evoluce (SNM), který je sám o sobě založen na dlouhé řadě teoretických argumentů, počínaje pracemi J.B.S. Haldana z 50. let minulého století. Chceme-li porozumět RAO, je důležité pochopit vývoj této teorie.
Haldane (1957)4 byl první, kdo se zabýval konceptem známým jako „náklady substituce“. Podle Haldana by přirozené populace neměly být schopny zvládnout počet úmrtí, který přírodní výběr vyžaduje k pozitivní evoluci. Jednoduše řečeno, vyšší obratlovci nemají dostatečně vysokou reprodukční rychlost, aby mohli podporovat rychlou evoluci prospěšných látek. K selekci nových mutací je zapotřebí příliš mnoho úmrtí („cena“). Mnozí kreacionisté tvrdí, že „Haldanovo dilema“ nebylo doposud dostatečně zodpovězeno.5
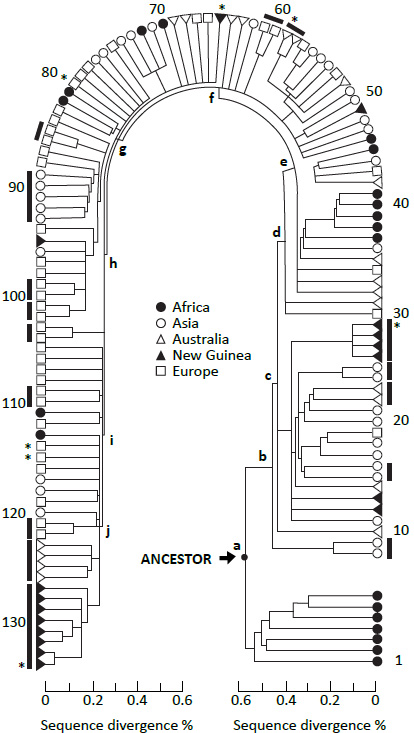
 Obrázek 2. Strom, kterým to všechno začalo. Cannová et al.6) založili svou „africkou“ teorii na skutečnosti, že první hlavní větev jejich stromu vede ke všem africkým sekvencím na jedné straně a smíšeným africko-světovým sekvencím na straně druhé. Tento závěr vychází z řady předpokladů, které jsou v textu podrobně rozebrány. (Viz obrázek 3, Cannová et al.7). Klikněte sem pro větší zobrazení. |
Motoo Kimura posunul Haldaneův argument o krok dále a aplikoval jej na měřitelné genetické rozdíly mezi savci. V návaznosti na Haldana usoudil, že pokud jsou všechny genetické rozdíly mezi dvěma druhy způsobeny pozitivním výběrem a pokud se vyvinuly v rámci standardního evolučního časového rámce, pak by savci potřebovali astronomicky vysokou míru reprodukce, aby měl přírodní výběr dostatek materiálu pro evoluční změny. Například lidé a šimpanzi mají miliony genetických rozdílů, ale za 3 miliony let bychom měli k dispozici pouze 100 000 lidských generací, během nichž bychom tyto miliony rozdílů napravili. Podle evolučních výpočtů by musel přírodní výběr odstranit mnohonásobně více lidí, než se jich v této době mohlo narodit, aby se napravilo tolik rozdílů. Kimura toto dilema vyřešil tak, že většina nových mutací musí být „neutrální“ (odtud pochází přesvědčení, že většina genomu je tvořena „odpadní DNA“; o pojmu se píše na jiném místě tohoto webu8). Rychlost neutrální evoluce by mohla být mnohem vyšší než pozitivní evoluce a byla by omezena pouze rychlostí chyb při kopírování DNA. Protože přírodní výběr nepůsobí na neutrální znaky, které neovlivňují přežití ani reprodukci, může neutrální evoluce probíhat náhodným driftem bez jakýchkoli vlastních nákladů na výběr. Kimura považoval Haldanovo dilema za závažný problém a uvedl ho jako hlavní důvod, proč navrhl neutrální evoluční teorii.9 Haldane měl pravdu, tvrdil Kimura, ale převaha biologických změn musí být neutrální.
Neutrální model byl mnoha autory rozšířen na tzv. standardní neutrální model (SNM). SNM byl vyvinut tak, aby pokryl složité historické vzorce, jako jsou události hrdla lahve. Jedná se o základní pilíř systému RAO.
Následuje seznam předpokladů, které jsou pro model RAO rozhodující. Všimněte si, jak SNM plyne ze samé podstaty RAO (obrázek 2):
Stálá velikost populace. Přestože neutrální model není ovlivněn velikostí populace, byl SNM vyvinut částečně pro modelování změn velikosti populace (konkrétně jednoho cyklu populačního hrdla lahve). RAO předpokládá jedinou lidskou populaci s jedinou expanzní událostí a bez následných subpopulačních hrdel lahve nebo jiných demografických rozdílů.
Náhodné páření (bez geografické či jiné substruktury populace). Neutrální teorie opět není podmíněna náhodným pářením, ale RAO ano. Substruktura populace před nebo po africké události by mohla skrývat skutečný obraz demografické historie lidstva.
Neutrální polymorfismy. Na dané alely nemůže působit žádná selekce. RAO vychází z předpokladu, že celý mitochondriální genom je v podstatě neutrální, nebo alespoň že negativní mutace jsou účinně eliminovány. Pro podrobnou kritiku této problematiky viz Sanford.10
Model mutace s nekonečným počtem míst. V různých liniích se nesmí vyskytovat více mutací na stejných místech a zpětné mutace nejsou povoleny. V jistém smyslu je to rozumná aproximace vzhledem ke značné velikosti genomu a malému počtu generací (v modelu Stvoření) nebo při nízké míře mutací (v evolučních modelech). Aproximace také zjednodušuje teoretické porozumění a výpočty. Pokud se však tato aproximace ukáže jako nesprávná, model RAO se stane hůře pochopitelným a výpočty, které za ním stojí, se stanou méně průkaznými.
Konstantní rychlost mutací / substitucí u všech subpopulací („molekulární hodiny“). Pro ilustraci, jak je to důležité, budu citovat Cannovou et al.:11 „Ke stromu lze připojit časové měřítko… za předpokladu, že divergence (odchylka) sekvence mtDNA se hromadí konstantní rychlostí“.
 Obrázek 3. Zjednodušený strom mtDNA z mitomap.org. Tento diagram ukazuje vztahy mezi hlavními mitochondriálními liniemi. Protože není prezentován v tradičním formátu stromu, je zřejmé, jak obtížné je určit, kam by měla být umístěna sekvence předků. Carter12 umístil kořen nikoli do Afriky, ale do písmene „R“, které se nachází blízko středu stromu. (Převzato z MITOMAP: A Human Mitochondrial Genome Database, mitomap.org, 2008). Klikněte sem pro větší zobrazení. |
Konstantní efektivní generační doba všech subpopulací (u lidí se obvykle předpokládá 20 let, ačkoli různí autoři používají hodnoty mezi 20 a 30 lety).
Společný předek člověka a šimpanze asi před 3-6 miliony let. Nejedná se o předpoklad SNM, je však nutný pro kalibraci události hrdla lahve SNM. Spočítáním počtu rozdílů mezi šimpanzími a lidskými mitochondriemi a vydělením tohoto počtu 3 až 6 miliony lze v podstatě získat odhad počtu mutací, které by se v populacích měly nahromadit za rok.
Tajimova statistika D
Tajimova statistika D13 se používá k testování SNM podél daného úseku DNA. Jedná se v podstatě o shrnutí spektra frekvencí alel. Hodnoty D, které se významně neliší od nuly, ukazují, že populace splňuje všechna kritéria pro SNM. Kladné a záporné hodnoty jsou způsobeny nadbytkem, resp. nedostatkem očekávaného počtu vzácných polymorfismů. Výrazné kladné nebo záporné hodnoty D mohou naznačovat přítomnost přirozeného výběru nebo historických změn ve velikosti populace či introgrese více populací (introgrese = vnesení genů jednoho druhu do genomu jiného mezidruhovým a následným zpětným křížením). To je důležité měřítko pro teorii SNM i RAO.
Očekává se, že historické změny ve velikosti populace ovlivní Tajimovu statistiku D v tom smyslu, že růst populace by měl vést k nadbytku nízkofrekvenčních polymorfismů (záporné hodnoty D). K tomu dochází proto, že nové mutace jsou přenášeny spolu s expanzí a z populace se tak snadno nedostanou náhodným driftem. Alternativně by populační hrdla lahve měla vést k nedostatku nízkofrekvenčních polymorfismů (pozitivní hodnoty D), protože většina nízkofrekvenčních alel je eliminována náhodnou selekcí.
Nerovnováha vazeb
SNM je třeba zasadit do kontextu pohlavního rozmnožování. Při tvorbě gamet dochází ke křížení mezi kopiemi chromozomů. Toto míchání způsobuje náhodnou změnu alel (variací) podél úseku DNA. Protože však v každé generaci dochází pouze k jednomu nebo dvěma křížením na jedno rameno chromozomu, nejsou v každé generaci randomizovány (náhodně seskládány) všechny alely. Čím blíže jsou si dvě alely, tím menší je pravděpodobnost, že budou odděleny. O alelách v těsné blízkosti se říká, že jsou „propojené“. To platí i pro alely oddělené oblastmi s řídkým křížením. Genetici používají termín „vazebná nerovnováha“ (LD) k popisu nerovnoměrného propojování určitých alel s určitými jinými alelami. Soubor společně děděných alel se označuje jako „haplotyp“.
Růst populace vede nejen k záporným hodnotám D (nadbytek alel s nízkou frekvencí), ale také ke snížení LD, protože křížením se v každé generaci náhodně získává stále více alel. Alternativně by populační hrdla lahve měla vést k vyšší LD, protože během hrdla lahve malý počet lidí předá své velké bloky vazeb celé populaci. Trvá nějakou dobu, než se haplotypy zakódují.
Očekávání SNM
Populační parametry používané při výpočtech SNM je třeba odhadnout, což není snadný úkol. Pro člověka je rychlost mutací založena na současné úrovni genetické rozmanitosti, předpokladu, že převážná většina této genetické rozmanitosti je neutrální, a předpokládaném společném předku šimpanze a člověka před 3-6 miliony let. Jinými slovy, výpočet předpokládá, že platí evoluce ve velkém měřítku (tj. makroevoluce). Efektivní velikost populace (Ne) se rovněž odhaduje na základě pozorovaných úrovní diverzity. Ale Ne závisí také na generační době a době, která nás dělí od společného předka, což jsou dva parametry, o nichž se toho ví jen málo.14 Za předpokladu stejného počtu samců a samic a náhodného páření činí poměr hypotetických Ne pro autozomy, chromozom X, nerekombinovatelnou část chromozomu Y (NRY) a mtDNA 4:3:1:1. Očekává se, že redukovaná Ne způsobí rychlejší diferenciaci mezi populacemi u haploidních lokusů než u ostatních.15 (Lokus je genetický termín označující přesné místo na chromosomu, na kterém se vyskytuje příslušný gen nebo polymorfismus. Konkrétní varianta genu či polymorfismu na příslušném lokusu se nazývá alela.) To je jeden z důvodů, proč byla RAO původně založena na mitochondriálních sekvencích.
„Hrdlo lahve spojené s Potopou by vytvořilo silný signál, který by měl být patrný ještě dnes. Jakýkoli model lidských dějin, který toto nebere v potaz, dojde k nesprávným závěrům, pokud je příběh o Potopě pravdivý.“
Protože výpočet doby k poslednímu společnému předku (TMRCA) je přímo úměrný Ne, očekává se, že TMRCA pro autozómy a chromozom X budou 4krát, resp. 3krát vyšší než pro oba haploidní lokusy.16 To však ignoruje možnost, že v zakládající lidské populaci byla pouze jedna mužská linie (Noemův chromozom Y) a pouze tři ženské linie (mitochondriální chromozomy tří Noemových snach). Hrdlo lahve spojené s Potopou by vytvořilo silný signál, který by měl být patrný ještě dnes. Jakýkoli model lidských dějin, který toto nebere v úvahu, dojde k nesprávným závěrům, pokud je příběh o Potopě pravdivý.
SNM předpokládá, že rozdíly v LD mezi populacemi jsou způsobeny rozdíly v demografické historii.17,18 Tento předpoklad samozřejmě závisí na konstantní míře rekombinace, mutace a genové konverze ve všech subpopulacích a na neexistenci staré populační struktury (všechny tyto faktory ovlivňují LD). Stačí však jediná změna v enzymu opravujícím DNA, jediná změna v genu, který ovlivňuje rychlost rekombinace, nebo jediná změna v genu, který ovlivňuje proces genové konverze v jedné z populací, a výsledky SNM se budou lišit od skutečnosti. SNM tedy předpokládá, že se populace v určitých genetických vlastnostech neodlišují, ale v jiných se mohou odlišovat. To je asi nejdůležitější bod, který je třeba pochopit.
Podle SNM by alely s nízkou frekvencí měly být obecně mladší. Jak se v populaci objevují nové alely, většina z nich se ztratí náhodným driftem. Nová alela má ve skutečnosti 1/Ne pravděpodobnost, že se nakonec stane fixní. Ve velkých stabilních populacích se očekává, že téměř všechny alely s nízkou frekvencí budou mladé. Mladší alely jsou také obecně spojeny s delšími haplotypy než alely s vysokou frekvencí,19 protože rozbití vazebných bloků a přemístění nových alel ve vztahu ke starším trvá dlouho. Předpokládá se, že rekombinace je neutrální a náhodná a že se vyskytuje s vyšší frekvencí než mutace.20
Podle definice platí, že čím více vazebných bloků populace má, tím větší je její očekávaná schopnost udržet polymorfismus (vazby zvyšují rozptyl počtu polymorfních míst21). Vyšší hladiny Ne také umožňují populaci udržovat polymorfismus (a vyšší počet vazebných bloků). Závěr, že africké populace si dlouhodobě udržují větší efektivní velikost populace než neafrické populace,22 je založen na úrovni polymorfismu zjištěné v populacích za předpokladů SNM. Jsou však tyto předpoklady platné?
Nesrovnalosti SNM
V evoluční literatuře byly zjištěny a publikovány nesrovnalosti, kterých se SNM dopouští ve všech svých předpokladech. Populace v reálném světě neodpovídají omezením SNM. Proto musíme být opatrní, když čteme závěry mnoha studií, které byly založeny na této evoluční teorii.
„Proto je třeba očekávat, že k nesrovnalostem SNM [standardního neutrálního modelu] bude docházet často. Lze oprávněně pochybovat o užitečnosti modelu, který nelze přizpůsobit reálným údajům.“
SNM je nezbytné zjednodušení, které umožňuje studovat velmi složitý systém. Nesrovnalosti SNM lze však očekávat na všech úrovních: struktura populace, hrdla lahve v subpopulacích, malé rozdíly v míře rekombinace a příměsi více populací zvyšují odhady LD23 a narušují propočty SNM. Vzorce haplotypů mohou být narušeny také opakovanými mutacemi, přeměnou genů, chybami při sestavování genomu a chybami v genotypování.24 Pokud dojde k odchylkám od SNM (např. závěry většiny studií provedených na evropské populaci), je význam odhadovaných populačních parametrů nejasný.25 V reálném světě se má za to, že se generace překrývají, že Ne v čase kolísá, že tok genů mezi populacemi se v čase mění a že v rámci populací po celém světě dochází k vytváření populační struktury. Proto je třeba očekávat, že k nesrovnalostem SNM bude docházet často. Lze oprávněně pochybovat o užitečnosti modelu, který nelze přizpůsobit reálným údajům.
Ne
K provedení většiny výpočtů SNM je třeba dobře odhadnout hodnotu Ne, avšak odhady Ne jsou ovlivněny několika parametry, jejichž vliv je často nutné odečíst, aby model mohl fungovat. Výpočty dlouhodobých hodnot Ne jsou neúměrně ovlivněny nízkými hodnotami (zmenšování populace):26 proto by uváděné hodnoty měly být spíše podhodnocené. Doba trvání generace také silně ovlivňuje Ne. Obecně se očekává, že delší generační doba bude mít za následek úměrně nižší Ne (a naopak). Většina studií LD předpokládá, že generační doby jsou u jednotlivých populací stejné, což nelze historicky prokázat. Kulturní i genetické rozdíly mohou vést k rozdílům v generační době. Generační doba odhadovaná z moderních genealogických údajů je větší než 20 let, které se v těchto studiích běžně předpokládají (například 10 generací v mém rodokmenu = 300 let, tedy 30 let/generace). A podle biblických údajů by neměla existovat konstantní generační doba, protože bezprostředně po Potopě se generační doba výrazně zkrátila.
Vyšší hladiny Ne by měly vést k menšímu genetickému driftu a zpomalit divergenci populací. Lidská Ne se často odhaduje na přibližně 10 000 lidí. Zajímavé je, že vzhledem k vypočtenému koeficientu příbuzenského křížení odhadují Reich et al.27 historickou lidskou Ne na „50 jedinců po 20 generací; 1 000 jedinců po 400 generací; nebo jakoukoli jinou kombinaci se stejným poměrem.“ Rád bych upozornil, že 5 jedinců na 2 generace odpovídá jejich poměru a zhruba odpovídá biblickým očekáváním. Je to důkaz Potopy (šest lidí na jednu generaci), nebo jsou tyto výpočty natolik zakořeněné v evoluční teorii, že nejsou užitečné? Frisse et al.28 zjistili nesoulad mezi odhady Ne pro neafrické populace na základě údajů o LD a polymorfismu. To je další příklad skutečných populačních dat, která jsou v rozporu s předpoklady SNM. Která Ne by se měla použít?
Z nejasných důvodů je odhad historické lidské Ne 10 000 jedinců mnohem nižší než Ne odhadů pro velké opice.29 Mezi žijícími šimpanzi existuje obrovská rozmanitost:30 možná třikrát až čtyřikrát větší než v celé lidské populaci.31 Podporuje tento důkaz evoluci podle scénáře SNM, nebo tato rozmanitost není ani tak důkazem dávné Ne, jako spíše důkazem rychle se zmenšujícího šimpanzího genomu?
Výpočet Ne předpokládá hluboký čas. V biblickém modelu se výpočty Ne neprovádějí, protože se nepředpokládá dlouhý věk. Spíše říkáme, že před přibližně 4500 lety (Potopa) došlo k populačnímu hrdlu lahve, kdy se světová populace zredukovala na 3 zakladatelské páry (Noemovi synové a snachy).
Nenáhodné křížení
„Předpokládá se, že v lidském genomu existuje rozsáhlá jemná variabilita frekvence rekombinace, a modely, které zahrnují rekombinační horká místa, jsou často lepší než modely, které předpokládají náhodnou rekombinaci.“
Protože toho o mechanismu, který řídí křížení, ani o frekvenci, s jakou k němu dochází, mnoho nevíme, často se předpokládá, že křížení je víceméně náhodné. To umožňuje snadnější výpočet LD. Tento jev však není zcela náhodný. Rekombinační „horká místa“ jsou známa již řadu let. Předpokládá se, že v lidském genomu existuje rozsáhlá jemná variabilita frekvence rekombinace, a modely, které zahrnují rekombinační horká místa, jsou často lepší než modely, které předpokládají náhodnou rekombinaci.32 Abychom lépe pochopili genetickou historii člověka, musíme lépe porozumět rekombinaci. V tomto tématu je mnoho prostoru pro zlepšení a mnoho naděje pro biblický model původu člověka. Samotná přítomnost dlouhých, nesmíšených vazebných bloků naznačuje mladý genom, ale potřebujeme více údajů.
Struktura populace
Dalším klíčovým předpokladem SNM je náhodné páření nebo neexistence populační struktury (konkrétně u subsaharských afrických populací před rozptýlením z Afriky). Očekává se, že efektivní rychlost rekombinace se ve strukturovaných populacích sníží, protože haplotypy omezené v rámci různých subpopulací nebudou mít možnost rekombinovat tak často jako v případě panmiktické populace (náhodně se pářící populace).33 Důležité je, že nerozpoznání struktury populace může vést k falešně pozitivním výsledkům při testování konstantní Ne.34 Při diskusi o možnosti populační struktury v předpokládané původní africké populaci se Garrigan a Hammer35 obávali, že struktura předků v populaci mohla mít za následek zvýšení Ne předků a tím zkreslila jejich výpočty. Behar et al.36 tvrdí, že raná evoluční historie člověka v Africe zahrnovala malé a izolované kmeny existující nezávisle po tisíce let. To je zásadní problém, protože u malých a izolovaných populací dochází k příbuzenskému křížení, rychlému driftu a rychlé akumulaci nových mutací. Tato situace odporuje základním předpokladům SNM a zároveň poskytuje potenciálně vynikající materiál pro podporu biblického modelu: pokud totiž několik subsaharských afrických populací existovalo po Potopě v takovém stavu, mohlo by to do značné míry vysvětlit, proč je mezi lidmi afrického původu větší genetická rozmanitost.
Model náhodného páření ignoruje realitu, protože struktura populace je faktem lidské existence.37 Jedinci mají tendenci vybírat si partnery ze stejných sociálních skupin38 a subpopulace v těsné blízkosti mohou být navzájem zcela izolovány. Studie Bulajevové et al.39 ukazuje na základě genealogické analýzy, že v jisté dagestánské (oblast Ruska při hranicích s Gruzií) vesnici s 2700 obyvateli bylo za devět generací uzavřeno pouze 10 manželství vesničanů s lidmi odjinud! Většina těchto sňatků byla přitom uzavřena s obyvateli sousedních vesnic. Bamshad et al.40 ukázali, že hinduistický kastovní systém dokládá významnou historickou událost – obrovskou invazi do Indie ze severozápadu. Muži a ženy z vyšších kast jsou geneticky podobnější Východoevropanům, zatímco muži a ženy z nižších kast jsou podobnější jihovýchodním Asiatům. Náboženský systém značné části indického obyvatelstva brání volnému míšení již tisíce let! To vše říká, že realistický model demografické historie lidstva by byl příliš složitý,41 zvláště pokud by používal špatný model lidských dějin. Vysoce komplexní model ovšem nemusí být nutný (např. Liu et al. 200642), ale přílišné zjednodušení je vždy nebezpečné.
Příměs obyvatelstva
Nejenže neplatí předpoklad neexistence populační struktury (jak celosvětově, tak v rámci dílčích populací), ale situace je ještě komplikovanější v důsledku mísení kdysi oddělených populací. Když se dříve oddělené populace opět spojí, heterozygotnost se zvýší. Míchání haplotypů, které vznikly odděleně, způsobuje zvýšení vypočtené LD, a to i na nespojitých místech.43 Příměs způsobuje značné rozdíly v genetickém původu jedinců v populaci.44 Zajímavé je, že blokové charakteristiky smíšené populace by se měly nejvíce podobat populacím s nejnižší LD.45 Jinými slovy, míšení může maskovat značné množství LD. Například fascinující příměs afrických žen z kmene Bantu a židovských mužů (předků kmenů Lemba v JV Africe) vytvořila populaci s neobvykle vysokým počtem dlouhých vazebných bloků (long-distance LD).46
Konverze genů
„Přestože je o genové konverzi známo jen málo, jedna nedávná studie dospěla k závěru, že modely využívající křížení a genovou konverzi odpovídají datům lépe než modely využívající pouze křížení.“
Jak známo, LD se snižuje v důsledku křížení; je ovšem možné, že působí i jiný, méně známý proces zvaný „genová konverze“. Jedná se v podstatě o proces, při kterém úsek DNA může být zkopírován na velmi podobný úsek jiné DNA v těsné blízkosti. Tento proces byl intenzivně studován u kvasinek, ale v případě savců je dosud poměrně neznámý.47,48,49 Předpokládá se, že genová konverze působí na krátké vzdálenosti a rozbíjí LD mezi úzce propojenými markery, kde je křížení méně pravděpodobné.50,51 Pokud je v modelu povolena genová konverze, výpočty Ne založené na parametru křížení se výrazně zmenší.52 Přestože je o genové konverzi známo jen málo, jedna nedávná studie dospěla k závěru, že modely využívající křížení a genovou konverzi odpovídají datům lépe než modely využívající pouze křížení.53 Možnost rekurentních mutací však tento obraz komplikuje tím, že teoreticky nadsazuje zdánlivou úroveň rekombinace a zkresluje míru genové konverze směrem nahoru.54 Navíc vysoká úroveň rekurentních mutací může fylogenetický signál zcela zneviditelnit.55
V důsledku genové konverze je termín „NRY“ (nerekombinující část lidského chromozomu Y) technicky nesprávný, protože v oblastech chromozomu Y specifických pro muže byla zjištěna rozsáhlá genová konverze (forma rekombinace).56,57 Očekává se, že vysoká úroveň genové konverze zpomalí rychlost divergence, protože systematicky eliminuje mutační události.
Ekvivalentní mutační rychlosti a aktivní selekce
Jedním z hlavních předpokladů SNM je, že ekvivalentní genetické sekvence v různých populacích se vyvíjejí podle principu molekulárních hodin. K tomu může dojít pouze tehdy, pokud na dané genetické sekvence nepůsobí selekce a pokud je rychlost mutací ve všech populacích stejná. Existují však důkazy, že molekulární hodiny nefungují v rámci příbuzných kladů (označení pro skupinu organismů, která zahrnuje společného předka a všechny z něho vzešlé potomky) africké mitochondriální haploskupiny L2. Dva ze čtyř kladů, které studovali Torroni et al.,58 byly „neúměrně odvozené“, takže dospěli k závěru, že jejich výsledky „nejsou v souladu s jednoduchým modelem neutrální evoluce s jednotnými molekulárními hodinami.“ Howell et al.59 naznačili, že u všech afrických mitochondriálních haploskupin L mohou existovat poruchy hodin a že existují rozdíly v rychlosti hodin mezi kódujícími a kontrolními oblastmi. Friedlaender et al.60 zašli tak daleko, že řekli, že proměnlivá rychlost mutací mezi mtDNA klady zpochybňuje užitečnost koalescenční61 statistiky a s ní spojených odhadů stáří.
To jsou klíčová zjištění, protože hluboce zakořeněné větve subsaharských linií jsou základem teorie RAO. Pokud mezi nimi nefungují molekulární hodiny, není možné africkou diasporu časově zařadit. Jediným důvodem pro tvrzení, že tyto linie pocházejí od předků, je to, že se více odlišují od zbytku světové populace… ale jak došlo k takové odlišnosti, by pak zůstalo otevřenou otázkou.
Jiné studie dospěly k závěru, že odchylky od SNM v Evropě a Asii mohou být způsobeny přirozeným výběrem. Přirozený výběr také nemusí být u všech kladů stejný, zejména proto, že klady nejsou rovnoměrně rozmístěny ve všech oblastech prostředí.62,63
Selekci mohou napodobovat i kulturní faktory. Čingischán je předkem překvapivého počtu lidí ve střední a východní Asii a snad až 0,5 % (1 z 200!) světové populace.64 Kdybychom z historických pramenů nevěděli o existenci Čingischána, jak by tato chybějící informace ovlivnila naše závěry o rozšíření asijských chromozomů Y? A pokud mohou mít takovéto nenáhodné události tak zásadní vliv na vzorce genetické diverzity, jak bychom mohli věřit molekulárním „hodinám“, které jsou tak silně závislé na náhodných mutacích a náhodném páření?
Závěry
„Pokud se africké sekvence nevyvíjejí podle principu molekulárních hodin, je třeba teorii nedávného afrického původu vážně přepracovat.“
Studie, na které se tento článek odvolává, poukazují na nejistou povahu teorie nedávného afrického původu. RAO se dalekosáhle opírá o standardní neutrální model evoluce, ale každý předpoklad SNM, a tedy i RAO, byl v evoluční literatuře otevřeně zpochybněn. Pokud se africké sekvence nevyvíjejí podle principu molekulárních hodin, je třeba teorii nedávného afrického původu vážně přepracovat. Totéž by platilo, kdyby selekce působila na neafrické sekvence. Pokud populace v reálném světě odporují hlavním předpokladům SNM, závěry studií, které předpokládají SNM, nejsou a nemohou být konečným slovem na toto téma.
Můžeme konstatovat, že předpoklady SNM nejsou zcela realistické. Skutečná situace je mnohem složitější, než umožňují zjednodušující předpoklady (obrázek 3), a některé z těchto předpokladů jsou buď zkreslené ve prospěch závěru, nebo v rozporu s údaji. Přechod není náhodný. Struktura populace existuje ve všech měřítkách. Příměs populací (zejména pokud k ní došlo v dávné minulosti) a genová konverze modely příliš komplikují. Přítomnost přírodního výběru mezi lidskými mitochondriemi odstraňuje „neutrální“ část SNM, pokud jde o mitochondriální Evu podle modelu RAO.
Konečně RAO a SNM vycházejí z přesvědčení, že evoluce může probíhat, ale že nemůže ovlivnit věci, které řídí rychlost evoluce. Nejsou možné rozdíly v mutačních rychlostech65,66 mezi populacemi způsobené stresem prostředí,67,68,69 demografií (vzorci životní dráhy způsobenými kulturními faktory) nebo mutacemi v DNA polymeráze a mechanismech kopírování, korektury nebo opravy DNA.70,71
Nyní se ukázalo, že nejpopulárnější evoluční model nedávné evoluce člověka je neuspokojivý. Čím jej však nahradíme? Biblický model genetické historie člověka je stále ještě v plenkách. Doufám, že tento krátký článek podnítí tvůrčí myšlení dalších vědců zabývajících se stvořením, kteří převezmou pochodeň a zaútočí na evoluční teorii v jejích kořenech, ale také, že se jim podaří představit nové myšlenky široké komunitě. Čeká nás mnoho práce. Zatím jsem jen nastínil několik myšlenkových směrů, které by mohly být pro kreacionistický výzkum docela přínosné.
Poděkování
Při psaní tohoto článku mě významně ovlivnil a částečně podpořil J. Sanford. Rád bych poděkoval zejména jednomu z recenzentů za vynikající kritiku, díky níž byl rukopis výrazně vylepšen.

{kind=link}
{kind=link}