
Kladistika, evoluce a fosilie
Nyní pracujeme na odborných a jazykových korekturách a na přípravě grafiky.
Kladistika je hlavní metodou používanou pro určování evolučních vztahů v biologii. Výsledky kladistických analýz, stromové diagramy zvané kladogramy, se často používají jako důkaz evoluce. Ačkoli byla kladistika vyvinuta evolucionisty a pro evolucionisty, stále nedokáže prokázat evoluci, natož biologickou realitu. Evoluce je obvykle stále považována za teoretické zdůvodnění používání kladistiky v paleontologii, takže evoluční závěr je pouze argumentací v kruhu. Kladogramy jen ukazují hierarchii biologických znaků, ale neříkají nic o tom, co tento vzorec vytvořilo. Evoluční kladistika také zobrazuje zjednodušený pohled na biologické změny a nezabývá se pleiotropií uvnitř organismů. Těchto problémů si byli někteří evolucionisté vědomi již před více než 30 lety, ale jejich kritika většinou zůstala bez odezvy, nejspíše proto, že jejich připomínky byly kreacionisty používány jako munice. Mnohé problémy fylogenetické inference, které má kladistika řešit, zůstávají stále z velké části nevyřešeny, například rozlišení mezi homologií a homoplazií. Největším problémem je však pravděpodobně iluze evoluce, kterou kladogramy a jazyk používaný k jejich popisu vytvářejí u veřejnosti. Obojí vytváří iluzi vyřešené genealogie navzdory tomu, že někteří kladisté odmítají jakékoli přísné genealogické konotace.
Co je kladistika?
Evoluční boje
Velká pozornost se pak soustředila na filozofické základy kladistiky. V 80. letech 20. století se většina systematiků shodla na tom, že kladistika je užitečná metoda. O tom, co kladistika znamená a k čemu má sloužit, se však vedly rozsáhlé debaty.
Dominantní myšlenková škola se odvíjela od Henniga a nadále tvrdila, že evoluce je nezbytným předpokladem pro fungování kladistiky.5 Hennigovi žáci důsledně nazývali kladistiku „fylogenetickou systematikou“ – pracovali s předpokladem, že evoluce je základem kladistiky. Proto se domnívali, že účelem kladistiky je objasnit nejpravděpodobnější evoluční vztahy, které se vyvíjely v průběhu historie. Kladistika se v podstatě stala cvičením v evolučním teoretizování.
| Komorové srdce |
Čelisti | Čtyři končetiny |
Plodové vejce |
Srst | Rození živých mláďat |
|
| Mihule mořská (Petromyzon marinus) |
1 | 0 | 0 | 0 | 0 | 0 |
| Pstruh duhový (Oncorhynchus mykiss) |
1 | 1 | 0 | 0 | 0 | 0 |
| Rosnice siná (Litoria caerulea) |
1 | 1 | 1 | 0 | 0 | 0 |
| Agama límcová (Chlamydosaurus kingii) |
1 | 1 | 1 | 1 | 0 | 0 |
| Ptakopysk podivný (Ornithorhynchus anatinus) |
1 | 1 | 1 | 1 | 1 | 0 |
| Kočka domácí (Felis catus) |
1 | 1 | 1 | 1 | 1 | 1 |
Tabulka 1. Jednoduchá kladistická analýza znaků, které jsou u obratlovců běžně považovány za společné odvozené znaky. Vlastnosti jsou polarizované: 0 – nepřítomnost a 1 – přítomnost.
Někteří systematikové však nesouhlasili s Hennigovým názorem, že kladistika musí nutně prokázat transformaci prostřednictvím změn znakového stavu.6 Tito takzvaní „transformovaní“ kladisté (kladisté „vzorců“) tento procesní předpoklad hennigovské kladistiky zpochybnili s tím, že jeho metodologie ke svému fungování nevyžaduje předpoklad evoluce. Pro transformované kladisty byla účelem kladistiky klasifikace založená na deskriptivní definici homologie. Kladistiku považovali za agnostickou, co se týče historie, a „evoluční historie“ založená na kladogramech nebyla ničím jiným než marnou spekulací. Ebach et al. to shrnují takto:
„Kladistika se nezabývá evolucí, ale vzorcem rozložení znaků v organismech nebo rozpoznáváním a charakterizací skupin.“7
Tento spor se přenesl do tehdejších sporů o „kreacionistickou vědu“.8 V této kontroverzi měly zvláštní vliv některé upřímné výroky Colina Pattersona, známého kritika hennigovské kladistiky.9 Odmítl spekulativní evoluční rekonstrukce, které mnozí systematici připojovali ke svým kladistickým analýzám, protože neexistoval způsob, jak ve skutečnosti identifikovat domnělé předky „označené“ uzly na kladogramu:
„S rozvojem kladistické teorie se ukázalo, že stále větší část evolučního rámce je nepodstatná a lze ji vynechat. Hlavním příznakem této změny je význam, který se v kladistice přikládá uzlům. V Hennigově knize, stejně jako ve všech zakladatelských pracích kladistiky, jsou uzly považovány za předky druhů. Tento předpoklad se ukázal jako zbytečný, dokonce zavádějící, a lze jej vynechat.“10
To vyvolalo poměrně relevantní otázku: proč se vůbec odvolávat na evoluci, když neexistuje způsob, jak rekonstruovat evoluční historii na základě morfologických srovnání? Je zcela v pořádku přiznat, že někdo věří v evoluci, ale transformovaní kladisté v podstatě zavrhli fosilní záznam jako její důkaz. Bez fosilního záznamu a biologického mechanismu evoluce zůstali transformovaní kladisté u zcela nevědeckého myšlení „evoluce mezer“. Není divu, že se to hennigovským kladistům nelíbilo; transformovaná kladistika potvrdila kreacionistickou kritiku evolučních „rekonstrukcí“ ze zkamenělin!
Ohlas
S rostoucí dostupností a výkonem počítačů v 90. letech minulého století se kladistika stala mnohem jednodušší, protože do té doby bylo porovnávání kladogramů s více než 15 znaky zahrnutými do analýzy nepřehledné.11 Velká část sporů pak utichla a Hennig byl v podstatě prohlášen za vítěze.9,12 Hennigovská kladistika se nakonec stala dominantní kladistickou metodou, kterou dnes systematikové používají, a to spíše na základě závazku akademické obce k evoluci než na základě skutečné závislosti kladistické metody na evoluci. Mnohé z důležitých otázek, které byly nastoleny v 70. a 80. letech, však zůstávají sporné i dnes, i když se o nich málokdy mluví tak otevřeně.
Neslučitelnost kladistických předpokladů a evoluce
Přestože kladistika byla původně určena k prokázání evoluce a nejpravděpodobnějších fylogenezí, existuje řada předpokladů, které jsou pro metodu kladistiky zásadní a které ji činí nevhodnou k prokázání evoluce.
Definice diskrétních, nezávislých proměnných v biologii
Kladistika považuje všechny znaky v rámci analýzy za diskrétní, nezávislé proměnné. Biologii však lze jen stěží popsat výhradně diskrétními pojmy; existuje mnoho vlastností živočichů, které jsou spojité. V biologii navíc existují složité vzájemné vztahy od molekulární po organismální úroveň, z nichž mnohým v současnosti nerozumíme. Proto může být definování znaku pro kladistickou analýzu i v genetice nesmírně obtížné. Tento problém je obecně výraznější u složitých morfologických prvků, jako jsou kosti, jak upozorňuje Lieberman:
„Kosti mají obecně nízký stupeň dědičnosti, protože jsou součástí složitých, integrovaných funkčních jednotek, které podléhají nejen mnoha genům s vícenásobnými účinky (pleiotropismus), ale také velkému množství negenetických vlivů. Proto je obtížné rozdělit kosti na samostatné, nezávislé jednotky fylogenetické informace. Z těchto důvodů mohou kosti a další aspekty morfologie přinést poměrně správné výsledky pro fylogenetické analýzy taxonomických jednotek vysoké úrovně, ale na nižších taxonomických úrovních, jako jsou druhy, jsou podstatně méně spolehlivé.“13
Jedním z hlavních důvodů tohoto problému je rozsah – čím menší jsou morfologické znaky použité pro analýzu, tím větší je obecně problém vzájemné závislosti. Výběr znaků se proto stává méně spolehlivým, stejně jako všechny interpretace homologie, které jsou na nich založeny. A protože homologie může ve skutečnosti existovat pouze na úrovni druhů, je prakticky nemožné prokázat evoluci pomocí kladistické metodologie, pokud od počátku nepředpokládáme evoluci.
„Společné odvozené vlastnosti“ a iluze linií
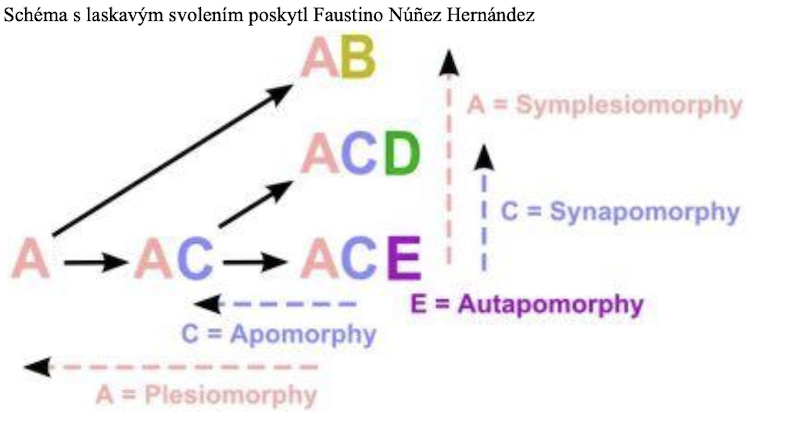
Synapomorfie neboli „sdílené odvozené znaky“ jsou charakteristické pro hennigovskou kladistiku.14 Jsou v kontrastu se sympleziomorfiemi neboli „sdílenými znaky předků“, které podle kladistů nemají pro kladistickou analýzu žádnou užitečnou informaci (obr. 2). Rozdíl mezi nimi je formálně nerozlišitelný, pokud se nepoužije postup zakořenění znaků, který udává směr kladogramu.15
V kladistice se synapomorfie obvykle ztotožňují s homologickými znaky, které jsou chápány jako důkaz společného původu.11,16 Jak bylo uvedeno výše, existují potíže s definováním „znaku“ v biologii způsobem užitečným pro kladistickou analýzu. Vezmeme-li však v úvahu tato omezení, můžeme na základě srovnání „společných“ znaků dospět k poměrně přesnému popisu morfologických vzorců podobnosti v rámci mnohobuněčného života.
Hlavní problém se sdílenými odvozenými znaky však spočívá ve slově „odvozený“. Podobnost ve formě nezaručuje společný původ, a to je interpretace kladogramu. Patterson zcela výstižně poznamenal:
„Přesto je těžké Gouldovi a lidem z Amerického muzea odporovat, když tvrdí, že žádné přechodné fosilie neexistují. … Řeknu to na rovinu – neexistuje jediná taková fosilie, pro kterou by bylo možné předložit neprůstřelný argument. Důvodem je to, že tvrzení o rodové příslušnosti a původu nejsou ve fosilním záznamu použitelná. Je Archaeopteryx předkem všech ptáků? Možná ano, možná ne: na tuto otázku nelze odpovědět. Je docela snadné vymyslet si příběh o tom, jak jedna forma vznikla z druhé, a najít důvody, proč by tyto fáze měly být zvýhodněny přírodním výběrem. Takové příběhy však do vědy nepatří, protože je nelze nijak ověřit.“17
„Kladistika neprokazuje evoluci. Kladistika je klasifikační schéma, nic víc.“
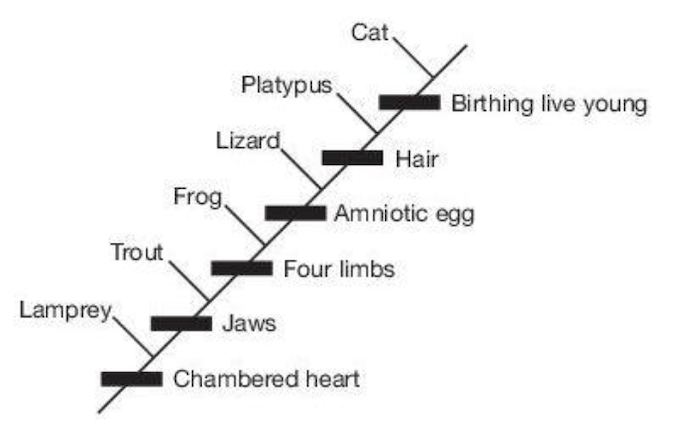
Všimněte si největšího problému: ze zkamenělin nelze zjistit, zda se zachovala nějaká linie. Kladistika neprokazuje evoluci. Kladistika je klasifikační schéma, nic víc. Woodmorappe to velmi dobře ilustruje (obrázek 3):
„Ale nedokazuje snad skutečnost, že se organismy dají uspořádat do vnořených hierarchií polarizovaných znaků (tj. kladogramů), že se takto (nebo vůbec) vyvinuly? Sotva. Za apriorního předpokladu evoluce by bylo možné sestavit kladogram, jehož korunovou skupinou je osmnáctikolový nákladní automobil a který ukazuje jasně vyplněný přechodný, postupný výskyt ‚nákladovosti‘, počínaje kmenovou skupinou jednokolky. Všimněte si také, že člověk, slon a netopýr jsou vysoce odvozené ryby, stejně jako osmnáctikolový náklaďák je vysoce odvozená jednokolka. To je reductio ad absurdum kladistické metodologie.“18
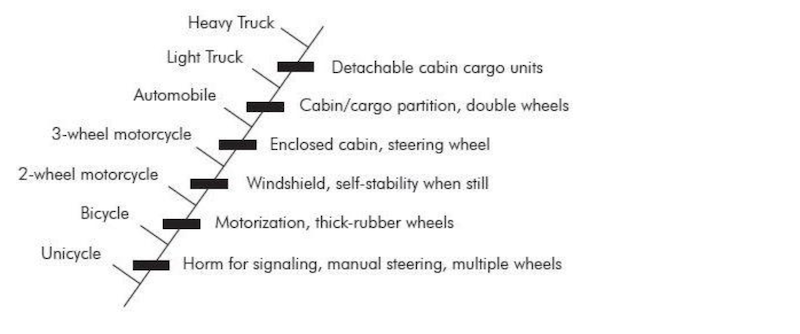
Není důvod předpokládat, že uzly kladogramu představují domnělé předky, a nejsou prokázány žádné přímé linie.19
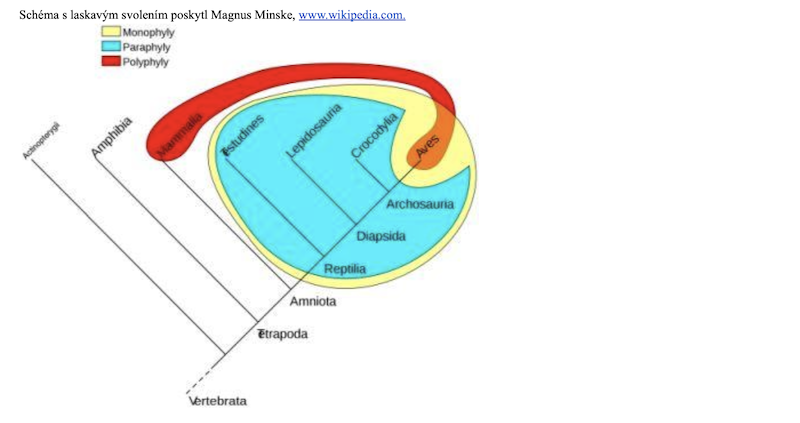
Co je to klad?
Klad je obvykle definován vztahy mezi předky a potomky; jedná se o předka a všechny jeho potomky. O všech taxonech v rámci kladů se říká, že jsou monofyletické, tj. obsahují všechny potomky pravděpodobně nejbližšího společného předka členů skupiny. Kladistika má být díky objevování synapomorfních znaků schopna rozlišit monofylii od parafylie (obr. 4). Parafyletická skupina je monofyletická skupina minus jeden klad v rámci této monofyletické skupiny. Příkladem jsou plazi, skupina, která v běžné řeči nezahrnuje ptáky, protože evolucionisté věří, že ptáci jsou údajně potomky plazů (teropodních dinosaurů). Endotermní živočichové (savci a ptáci) by byli příkladem polyfyletické skupiny, protože endotermie je podle evoluce homoplastický znak.
Opět však narážíme na problém, že metodika kladistiky neprokazuje rodokmen. Předek je v paleontologii nejen neznámý, ale i nepoznatelný. Evoluční rozsah kladů je však také libovolný: může jít o cokoli od jediného rodiče a dcery až po celou biologii (protože ortodoxní evoluce předpokládá, že vše vzniklo z jednoho společného předka, nikoli z mnoha). Kladisté tvrdí, že všechny ostatní klasifikační metody jsou z biologického hlediska nepoužitelné, protože genealogie je jediným prostředkem, jak prokázat relativní podobnost mezi různými organismy. Ernst Mayr však upozorňuje, že kladisté plně nedoceňují genetické a fenotypové vzdálenosti, které mohou existovat mezi domnělými „sesterskými taxony“:
„Jedna z několika fyletických sesterských linií může vstoupit do nové adaptační zóny a tam být vystavena silným novým selekčním tlakům. V důsledku toho se bude dramaticky lišit od svých kladisticky nejbližších příbuzných a může se geneticky natolik odlišit, že by bylo biologicky zavádějící nazývat sesterské skupiny blízkými příbuznými. Protože jsou však společnými potomky jednoho kmenového druhu, musí být označeny jako sesterské skupiny. A protože se jedná o sesterské skupiny, musí být jejich postavení koordinované, tj. podle kladistické teorie musí mít stejné absolutní kategoriální postavení v hierarchii (Hennig 1966, 139). Toto rozhodnutí ignoruje skutečnost, že jeden z nich je stále velmi podobný kmenovému druhu, zatímco druhý se mezitím vyvinul v naprosto odlišný typ organismu.“20
Mayr také rozlišuje mezi různými typy divergence, které kladistika nedokáže identifikovat: „Jakákoli teorie klasifikace, která nevěnuje pozornost obrovskému rozsahu rozdílů mezi posuny fyletických linií do menších nik a do zcela nových adaptačních zón, musí nutně vést k nevyváženým a nesmyslným klasifikacím. Takové zanedbávání různých druhů fyletického vývoje je však přesně to, co vyžaduje kladistická metoda.“21
Kladistika uznává pouze změny, které vznikají v důsledku větvení (kladogeneze), a tím homogenizuje různé typy morfologických rozdílů, což vytváří biologicky nereálnou situaci. Ironií osudu se tak fyletičtí gradualisté (jako Mayr) stávají nevědomými spojenci kreacionistů, protože i my uznáváme různé typy morfologických rozdílů. Rozdíl je samozřejmě v tom, že Mayr et al. jsou (navzdory důkazům) přesvědčeni, že velkou morfologickou diskontinuitu lze překlenout naturalisticky – kreacionisté nikoli. Nicméně tato neschopnost rozlišovat mezi různými typy biologických rozdílů činí kladistiku nevhodnou pro rekonstrukci historie biologie.
Evoluce nevyžaduje vnořený vzor
Kladistika předpokládá, že její srovnávací jednotky lze uspořádat do vnořené hierarchie.22 Evolucionisté předpokládají, že evoluce je jediným schůdným vysvětlením pro vzorec vnořené hierarchie. Hennigovci jdou ještě dál a tvrdí, že to z evoluce činí životaschopnou procesní teorii, která dává kladistice reálný smysl a ospravedlňuje její použití v systematice.23 To však prokazatelně není pravda. Vzorce vnořené hierarchie v přírodě nejsou závislé na evolučních předpokladech, protože byly rozpoznány mnohem dříve, než byla vědeckou komunitou přijata naturalistická evoluce:
„Ačkoli není v zásadě prokazatelná z vnějších důkazů (Panchen, 1992), existenci jediné, nepravidelně se větvící hierarchie vztahů mezi biologickými taxony považuje Brady (1985) za empirický fakt na základě jejího historického vzniku jako převládajícího prostředku k reprezentaci vzorců taxonomického seskupování, který používali předevoluční systematikové na počátku 19. století. To, že se tak stalo ještě před všeobecným přijetím evoluční teorie vědeckou komunitou, je jasným důkazem, že hierarchické pojetí přírodního systému není závislé na teorii evolučních procesů (Crow, 1926; Platnick, 1982).“24
Pokud evoluce nebyla nutná k pojetí života jako vnořeného vzorce, pak je vnořený vzorec života přizpůsoben evoluci, nikoli evolucí předpovězen nebo potvrzen. Když se Hennig snaží stanovit teoretickou prioritu evoluce na vnořené hierarchii,23 nevidí svůj anachronický a nedostatečně podložený předpoklad naturalismu. Darwin vycházel z vnořeného uspořádání života, které již bylo prokázáno nezávisle na evoluci. Poté vytvořil jednoznačně naturalistické vysvětlení jeho vzniku.
Evoluce však nevyžaduje vnořený vzor, protože se stejně snadno, ne-li lépe, může přizpůsobit jiným vzorům.25 Například transpozice (známá také jako laterální přenos genů) by poskytla mnohem rychlejší mechanismus šíření nových genů/struktur v biosféře než společný původ. Evolucionisté by stále předpokládali, že došlo k původu s modifikací, protože poskytuje mechanismus biologické novosti. Rozsáhlá transpozice by však do jakéhokoli vnořeného vzoru, o němž se předpokládá, že odpovídá původu s modifikací, vnesla tolik šumu, že by se vnořený vzor ztratil. Evolucionisté neuznávají transpozici jako rozšířený jev, zejména u mnohobuněčného života, jen proto, že nejsou pozorovány vzory, které by transpozici naznačovaly.
Navíc ani společný původ nevyžaduje vnořený vzor.26 Protože se předpokládá, že znaky mají nezávislou fylogenezi a rychlost evoluce, není zaručeno, že i blízké sesterské taxony budou mít relativně podobnou morfologii ve srovnání se vzdálenějšími příbuznými organismy. Kromě toho transformace v rámci linie (anageneze) nevytváří vnořený vzor, protože transformace, ke které mělo dojít, nebyla způsobena událostí větvení. Homoplazie tuto otázku ještě více komplikuje, protože může způsobit, že vzdáleně příbuzní tvorové jsou si morfologicky podobnější než domnělé sesterské taxony. Společný původ má přístup ke skutečnému balíku vysvětlení, která nemusí vytvářet vnořený vzorec.
Zastánci kladistiky vzorců sice odmítají evoluci jako teoretické zdůvodnění kladistiky, ale přesto věří, že je to jediné možné vysvětlení. Běžná konstrukce však vysvětluje i takový vzorec, a to s potenciálně větší silou.27 Pokud je život navržen tak, aby vyslal robustní zprávu, že je produktem jednoho designéra, vnořená hierarchie tuto úlohu splní. I když příjemce zprávy (my) má značně neúplné pochopení dat (kvůli vymírání druhů nebo neschopnosti prozkoumat všechna data), vnořený vzorec sjednocuje život, je plný homoplazií a také představuje dostatečně velké morfologické mezery mezi různými formami života, aby znemožnil společný původ. Život tak vysílá jednotnou, nikoli naturalistickou zprávu: je produktem jednoho designéra, který život navrhl tak, aby odolal naturalistickým vysvětlením jeho vzniku.
„Ani evoluce obecně, a zejména ani původ s modifikací nevyžadují vnořený vzorec. Vnořený vzorec lze navíc přinejmenším stejně dobře vysvětlit v běžném paradigmatu designu.“
Kladistika vyžaduje vnořený vzorec a fosilní důkazy do něj poměrně dobře zapadají, zejména u vyšších taxonomických kategorií. Nicméně ani evoluce obecně, a zejména ani původ s modifikací nevyžadují vnořený vzorec. Vnořený vzorec lze navíc přinejmenším stejně dobře vysvětlit v běžném paradigmatu designu. Proto si evoluce nemůže nárokovat, že je logickým zdůvodněním kladistiky, a není jediným dostupným vysvětlením takového vnořeného vzorce. Ani evolucionisté nemohou oprávněně považovat kladistiku za přesný odraz skutečné fylogeneze, protože evoluce vyžaduje anagenezi, nejen kladogenezi.
Problémy s výsledky a interpretací
Homologie
Problémy, které koncept homologie představuje pro evoluci obecně, byly dobře zdokumentovány jinde a nebudeme se k nim zde vracet.28 Je však třeba uvést několik důležitých poznámek týkajících se homologie a kladistické metody.
Kladistika jako metodologie může pomoci identifikovat homologii v závislosti na použité definici homologie. Definování homologie s ohledem na kladistickou analýzu se ukázalo stejně obtížné jako u jiných systematických metod.16 Termín „homologie“ pochází od Richarda Owena a ten ji chápal pouze jako podobné struktury používané pro různé funkce. Darwin definoval homologii podobným způsobem:29
„Všichni fyziologové připouštějí, že plovací měchýř je svou polohou a stavbou homologický nebo ‚ideálně podobný“ plicím vyšších obratlovců…“
Zbytek věty ukazuje, že homologii chápe jako podporu společného původu:
„… proto se mi zdá, že není velký problém věřit, že přírodní výběr skutečně přeměnil plovací měchýř na plíce nebo orgán sloužící výhradně k dýchání.“
Teprve postdarwinističtí biologové definovali homologii jako „podobné struktury vzniklé společným původem“. Definovali vysvětlení společného designéra mimo existenci.
Někteří transformovaní kladisté si tento rozdíl uvědomili a od té doby opustili „tradiční“ evoluční definici homologie a přijali něco bližšího Owenově deskriptivní definici.30 Homologie tedy může, ale nemusí prokazovat společný původ, ale společný původ je pro kladistický vztah irelevantní, protože společný původ se stává spíše historickým vysvětlením homologie než homologií z definice.
Homoplazie
Jednou z největších otázek, s nimiž se evolucionisté potýkají při morfologických analýzách, je, jak odlišit homoplazii od homologie. Homoplastické znaky jsou si podobné funkcí, ale mají odlišné základní struktury, a proto je nelze vysvětlit společným původem. Zřejmým příkladem jsou různé typy křídel: křídla hmyzu, ptáků, netopýrů a ptakoještěrů mají velmi odlišnou strukturu, ale stejnou funkci – létání. I ti nejzarytější obhájci hennigovské kladistiky považují za samozřejmé, že homoplazie je v kladistických analýzách běžná.31 Homoplazie vytváří šum v každém kladogramu, protože může vést k falešné identifikaci homologie, pokud není správně identifikována. Problémem se však stává měřítko – čím menší jsou části, na které znaky rozdělíme, abychom jich získali více, tím subjektivnější bude výběr znaků. Woodmorappe na to upozorňuje:
„… zatímco vnořená hierarchie může dobře charakterizovat živé organismy, pokud se na ni pohlíží z hlediska obecných podobností a rozdílů, neexistuje, pokud se současně bere v úvahu velké množství detailních morfologických podobností a rozdílů.“32
Jelikož se navíc ukázalo, že mnohé struktury, které byly na morfologické úrovni považovány za homologní, jsou na molekulární a/nebo vývojové úrovni homoplastické,28 argument homologie se ještě více oslabuje.
Mozaiková evoluce
Pro evolucionisty je důsledkem definování morfologických znaků jako nezávislých proměnných to, že jednotlivé znaky mají nezávislou fylogenezi, jinak známou jako mozaiková evoluce. Jednotlivé znaky se mohou evolučně zrychlovat, zpomalovat, zastavovat nebo obracet, a to nezávisle na ostatních znacích. Pokud se však znaky mohou takto vyvíjet, proč bychom měli očekávat vnořený vzorec na rozdíl od jakéhokoli jiného? Takový koncept může vysvětlit cokoli, což ho činí nefalzifikovatelným.
To také znamená, že organismy, které mají kombinaci plně vytvořených znaků vyskytujících se v různých kladech, se nazývají „střední“ nebo „přechodné“ fosilie. Takové tvory je však lepší označovat jako „mozaiky“ a byli plně funkční. Navíc nebyla prokázána žádná transformace, pouze například ryby s některými znaky čtyřnožců (Tiktaalik)33 nebo ptáci s některými znaky plazů (Archaeopteryx).34
Mozaiková evoluce je v podstatě všudypřítomná homoplazie bez zřetelného evolučního vzorce. Má tedy neomezený vysvětlující rozsah, ale za vysokou cenu obětování veškeré vysvětlující síly. Evoluce potřebuje empiricky prokazatelný mechanismus, aby byla historicky věrohodná, a žádný životaschopný mechanismus nebyl nikdy prokázán a ani není pravděpodobné, že bude.35,36 Mozaiková evoluce však tento problém stonásobně zhoršuje. Všechny diskontinuity a zvraty, které má mozaiková evoluce vysvětlit, je třeba vysvětlit mechanicky. Takové mechanistické vysvětlení by však bylo beznadějně složité a rozporuplné, protože by muselo vysvětlovat všechny možné evoluční scénáře současně. Mozaiková evoluce je tedy jen kouřovou clonou, která zakrývá skutečnost, že mechanistické vysvětlení fosilního vzorce, který popisuje, by bylo beznadějně složité a rozporuplné, nemluvě o biologické nereálnosti.
Člověk si pak klade otázku: pokud by tomu tak bylo, použili by evolucionisté někdy tak směšné vysvětlení fosilních vzorců? U většiny hlavních evolučních řad obratlovců: například v evoluci čtyřnožců,37 ptáků,38,39 savců40 a velryb,41 bylo zjištěno mnoho diskontinuit a zvratů v jednotlivých povahových stavech. Daeschler et al., uprostřed všech fanfár kolem objevu dnes již slavné „přechodné fosilie“ Tiktaalik, popisují sérii fosilií od ryb k čtyřnožcům (včetně Tiktaalika) tímto způsobem:
„Hlavní prvky tělesného plánu tetrapodů vznikly jako posloupnost přechodných morfologií, které se mozaikovitě a paralelně vyvíjely v třídě nozdratých, která je blízce příbuzná tetrapodům, což jí umožnilo využívat různá prostředí v devonu [zvýraznění přidáno].“42
Evolucionisté jsou pak nuceni odvolávat se na mozaikový vývoj kvůli četným diskontinuitám a zvratům, které se v řadě vyskytují. Tento vzorec je přítomen u většiny hlavních fosilních „evolučních přechodů“ obratlovců, a to navzdory 150 letům hledání nesčetných přechodných forem, které předpověděl Darwin. Pokud musí evoluce při vysvětlování fosilních vzorců tak silně spoléhat na mozaikovou evoluci, pak evoluce prostě nemůže vysvětlit vzorce ve fosilním záznamu.
Důraz na morfologii na úkor časové osy
Kladistika se zaměřuje především na morfologii a pracuje s vlastní idealizovanou časovou osou, která se řídí kladogenezí. To je nominálně v pořádku pro porovnání existujících tvorů, protože neexistuje žádná samostatná srovnávací časová osa. Pro evoluční paleontologii je to však problematické, protože často dochází ke konfliktům s datováním fosilií a idealizovanou morfologickou „časovou osou“, kterou vytváří kladogram. Výsledkem je, že mnoho morfologických analýz vede k „paradoxu dědečka“, kdy organismy, které jsou podle kladistické analýzy považovány za „předky“, jsou ve skutečnosti podle evolucionistů o miliony let mladší než jejich údajní potomci. Nedávným příkladem jsou stopy tetrapodů z Polska, které byly „datovány“ do doby o 20 Ma dřívější než Tiktaalik, běžně uváděný jako „přechodná fosilie“ mezi rybami a tetrapodem.43 Často se jim také říká „linie duchů“, protože tyto údajné organismy „předků“ nezanechávají ve zkamenělinách žádné stopy tam, kde bychom je očekávali.
Někdy je rozdíl „jen“ několik milionů let, což činí fosilie náchylnými k „redataci“, protože několik milionů sem nebo tam zastánci dlouhého věku obvykle neřeší. Jednou z oblastí, kde to představuje velký problém, jsou však ortodoxní spekulace o linii „od dinosaurů k ptákům“. Evolucionista Peter Dodson tento problém pěkně shrnuje:
„Osobně považuji i nadále za problematické, že ptákům nejpodobnější maniraptorní teropodi se objevují 25 až 75 milionů let po vzniku ptáků… „Linie duchů“ jsou upřímně řečeno umělým řešením, deus ex machina, které vyžaduje kladistická metoda. Samozřejmě se připouští, že maniraptoři svrchní křídy nejsou skutečnými předky ptáků, pouze ‚sesterskými taxony‘. Máme snad věřit, že skupina vysoce odvozených, rychle se vyvíjejících maniraptorů v jurském období dala vzniknout ptákům, jak vidíme u Archeopteryxe, a tato vysoce progresivní linie se pak dostala do stavu evoluční stáze a přetrvala v nezměněných základních rysech po miliony let? Nebo jsou skuteční předkové morfologicky mnohem bazálnější a hůře zařaditelní? Pokud ano, proč trvat na tom, že problém je vyřešen?“44
Co se týče evoluce „od dinosaurů k ptákům“, je ironií, že tento problém je snad nejhorší u nejbazálnějšího v současnosti známého dromaeosaurida (který je údajně nejbližším dinosauřím příbuzným ptáků), Mahakala omnogovae.45 Dochované fosilie Mahakaly jsou „datovány“ do doby před 80 Ma, ale k rozdělení mezi dromaeosauridae a paraves údajně došlo asi před 140 Ma.46 Navíc existuje mnoho dromaeosaurů, kteří tuto chronologickou mezeru vyplňují, ale všichni jsou morfologicky „pokročilejší“ než Mahakala. Jedná se o linii duchů, která vznikla před 60 Ma!
Kladogramy a iluze evoluce
ReMine označuje kladogramy za jednoho z hlavních viníků iluze evoluce.47 Evoluční strom je mocný obraz, který byl jedním z charakteristických znaků veřejného obrazu evoluce, a kladistika využívá stejné symboliky, protože kladogramy vypadají velmi podobně jako tradiční evoluční stromy. Čerpá svou sílu z toho, že údajně dokládá „linii“, kterou veřejnost automaticky interpretuje jako něco podobného rodokmenu (obrázek 1).48 Zastánci kladistiky vzorců často pohotově zdůrazňují, že kladogramy nejsou rodokmeny v pravém slova smyslu, ale že mají být nejlepšími odhady modelů cesty, kterou se evoluce ubírala. Hennigovští evolucionisté zároveň rádi opakují mantru, že kladistika „je nejčistší ze všech genealogických systémů klasifikace, protože pracuje pouze s blízkostí společných předků v čase [důraz přidán]“.8
„Kdyby veřejnost skutečně pochopila, co mají kladisté na mysli, evoluce by pravděpodobně ztratila velkou část důvěry veřejnosti, protože by bylo zřejmé, že nelze prokázat základní podmínku Darwinovy teorie: původ s modifikací.“
Učebnice kladistiky mohou být plné genealogických termínů, jako by kladistika a genealogie mluvily o tomtéž.49 Slova jako „rodový“, „odvozený“, „rodokmen“, „genealogie“, „primitivní“, „pokročilý“ atd. se neustále používají k tomu, aby veřejnosti přiblížila vztahy mezi taxony zjištěné kladistikou. Kladistika však nikdy neidentifikuje předky: používá nesčetné množství jiných metod, aby ukázala, jak mohl předek vypadat.15 To působí zmatek, protože kladogramy vzbuzují dojem, jako by byly podobnou demonstrací genealogie jako rodinné historie. Proto bude veřejné používání termínů s genealogickými konotacemi vždy uvádět v omyl většinu veřejnosti, která zná jen málo složitostí biologické systematiky. Zdá se, že existuje jediné schůdné řešení, jak se těmto nejasnostem vyhnout: přestat tyto termíny používat.
Domnívám se však, že tato upřímnost by si nevyhnutelně vyžádala vysokou cenu. Kdyby veřejnost skutečně pochopila, co mají kladisté na mysli, evoluce by pravděpodobně ztratila velkou část důvěry veřejnosti, protože by bylo zřejmé, že nelze prokázat základní podmínku Darwinovy teorie: původ s modifikací.
Závěry
Zkameněliny jsou vrtkavé. Jsou fragmentární, vzácné a umožňují protichůdné a rozporuplné interpretace. Navíc se často ukázalo, že kladogramy založené na morfologii jsou v naprostém rozporu s embryologickými a molekulárními daty. Když se pak kombinuje kladistika a analýza fosilií, vzniká beznadějně subjektivní hra na evoluční teorii, která nemá žádnou moc nezávisle prokázat evoluci. Tato subjektivita se ještě zhoršuje, protože kladistické analýzy jsou často v naprostém rozporu s datováním fosilií. Pro sladění časové osy implicitně obsažené v kladogramu s uznávanou časovou osou fosilií jsou obvykle nutné hypotézy ad hoc.
Tím, že kladistika považuje kladogenezi za jedinou metodu změny stavu znaku, ignoruje různé typy biologických rozdílů. Jednoduše extrapoluje známé mechanismy speciace a předpokládá, že mohou vytvářet komplexní novinky, což kreacionisté i mnozí evolucionisté odmítají. Kromě toho je definování znaků pro kladistickou analýzu ošemetné, protože vzájemně závislé znaky mohou zkreslovat analýzy, což se zhoršuje při výběru znaků s vyšším rozlišením. Tím, že kladistika v podstatě „digitalizuje“ taxony a linearizuje biologické rozdíly, vytváří biologicky nereálnou situaci a málo vypovídá o pravdivosti či nepravdivosti evoluce nebo stvoření.
Nicméně se kladogramy vydávají za důkaz evoluce, a přesto nedokážou identifikovat předky a potomky. Jazyk označující předky a používání kladogramů při prezentaci fylogeneze se často používá k vyjádření toho, co systematikové chápou jako pouhou topologii. „Evoluční strom“ je již 150 let mocnou metaforou používanou k demonstraci evoluce a kladogramy využívají tento obraz v povědomí veřejnosti, ať už to odborníci zamýšlejí, nebo ne. Pro veřejnost je to matoucí, protože špatně chápe, co kladogramy vlastně ukazují.
Kladistika nám umožňuje získat představu o hierarchii života, ale sama metoda nám neříká nic o tom, co tento vzorec vytvořilo. Protože evoluce nebyla nutná ani k objevení, ani k pochopení tohoto vzorce, nejedná se o důkaz evoluce. Kreacionisté se proto nemusí obávat toho, co se snaží kladistika ukázat. Nicméně to také zdůrazňuje potřebu vhodné systematické metody, která by mohla prokázat biologické rozdíly. Dokud se kreacionisté a evolucionisté nenaučí komunikovat pomocí systematiky, která dokáže empiricky identifikovat biologické rozdíly, dorozumění nikdy nebude možné.
Ironií je, že kladistiku vyvinuli evolucionisté pro evolucionisty, a přesto se jí nedaří prokázat evoluci, natož biologickou realitu. To naznačuje, že problém nespočívá ani tak v metodě, ale v základní teorii, kterou se pokouší demonstrovat. Kladogramy jsou modely vzorců života a jako takové mají svá omezení. Vykládáme-li však evoluci, která je nevyhnutelně genealogická, pomocí metody, která se výslovně vyhýbá pojmům genealogie, bude se zdát, že evoluce utíká před realitou.
